|

Рефератыполиграфия и этнографии москвоведению и спорту языку |
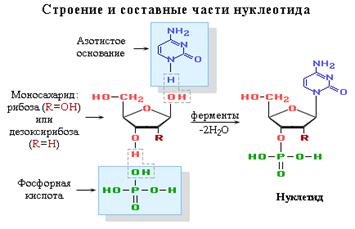
Реферат: Біохімія нуклеїнових кислотРеферат: Біохімія нуклеїнових кислотБіохімія нуклеїнових кислот ПланВступ. 3 І. Нуклеїнові кислоти. 4 1.1 Склад нуклеїнових кислот. 4 1.2 Значення нуклеїнових кислот. 6 ІІ. ДНК.. 8 2.1 Склад ДНК.. 8 2.2 Макромолекулярна структура ДНК.. 9 ІІІ. РНК.. 12 3.1 Склад РНК.. 12 3.2 Макромолекулярна структура РНК.. 12 Список використаної літератури. 15 ВступМи народжуємося, дорослішаємо, у нас з'являються діти і внуки. Ми жодн живі істоти на цій планеті, навколо нас щомить відбувається зародження нового життя. Цей процес не уривається ніколи. Наші сусіди по планеті – це мільярди живих істот: рослини, тваринні, мікроорганізми, віруси. Нас радує квітучий вишневий сад і шерех жовтіючого, відмираючого листя під ногами, утихомирює виплигуючі з води дельфіни стрибаюча білка – летяга. Всі ми коли-небудь хворіли на грип, краснухою і ц хвороби викликані знаходженням в нашому організмі хвороботворних мікробів і вірусів, а це теж живі організми. Як рідко ми замислюємося, звідки така різноманітність життя, і її форм, не таких схожих один на одного! А тим часом всі жив організми складаються з одних і тих же хімічних елементів, об'єднаних в макромолекули, такі як білки. Тільки у різних живих істот білки різні по своїй структурі. Але чому клітки певного організму синтезують тільки властиві їм білки? Як відбувається механізм передачі спадкової інформації, а головне – де вона зберігається? І. Нуклеїнові кислоти1.1 Склад нуклеїнових кислотНуклеїнові кислоти - це біополімери, макромолекули яких складаються з ланок, що багато разів повторюються, - нуклеотидів. Тому їх називають також полінуклеотидами. Найважливішою характеристикою нуклеїнових кислот є їх нуклеотидний склад. До складу нуклеотіда - структурної ланки нуклеїнових кислот - входять три складові частини: азотна основа - пірімідінове або пурінове. У нуклеїнових кислотах містяться підстави 4-х різних видів: два з них відносяться до класу пуринів два – до класу пірімідінів. Азот, що міститься в кільцях, додає молекулам основні властивості. моносахарид - рібоза або 2-дезоксирібоза. Цукор, що входить до складу нуклеотиду, містить п'ять вуглецевих атомів, тобто є пентозу. Залежно від виду пентози, присутньої в нуклеотиді, розрізняють два види нуклеїнових кислот рібонуклєїнові кислоти (РНК), які містять рібозу, і дезоксирибонуклеїнов кислоти (ДНК), що містять дезоксирібозу. залишок фосфорної кислоти.
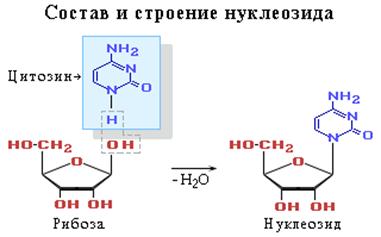
Нуклеїнові кислоти є кислотами тому, що в їх молекулах міститься фосфорна кислота. Нуклеотід - фосфорний ефір нуклеозіда. До складу нуклеозіда входять два компоненти: моносахарид (рібоза або дезоксирібоза) і азотна основа.
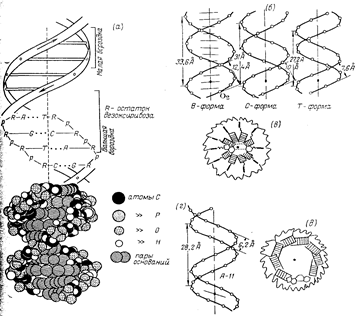
В кінці 40-х - початку 50-х років, коли з'явилися такі методи дослідження, як хроматографія на папері і УФ-СПЕКТРОСЬКОПІЯ, були проведен численні дослідження нуклеотидного складу НК (Чаргафф, А.Н. Белозерській). Одержан дані дозволили рішуче відкинути старі уявлення про нуклеїнові кислоти, як про полімери, що містять тетрануклеотидні послідовності (так звана тетрануклеотидна теорія будови ПК, пануюча в 30-40-е роки), що повторюються, і підготували грунт для створення сучасних уявлень не тільки про первинну структуру ДНК і РНК, але про їх макромолекулярну структуру і функції. Метод визначення складу ПК заснований на аналізі гидролізатів, що утворюються при їх ферментативному або хімічному розщеплюванні. Звичайно використовуються три способи хімічного розщеплювання НК. Кислотний гідроліз в жорстких умовах (70% -ва хлорна кислота, 100°С, 1ч або 100% -ва мурашина кислота, 175 °С, 2 ч), вживаний для аналізу як ДНК, так і РНК, призводить до розриву всіх N-гликозидних зв'язків утворення суміші пурінових і пірімідінових основ. При дослідженні РНК можуть використовуватися як м'який кислотний гідроліз (1 н. соляна кислота, 1OO°C, 1 ч), в результаті якого утворюються пурінові підстави і пірамідіпові нуклеозід-2'(3') - фосфати, так і лужний гідроліз (0,3 н. їдкий натрій, 37 °С,20 ч), що да суміш нуклеозід - 2' (3') - фосфатів. Оскільки в НК число нуклеотидів кожного вигляду рівно числу відповідних основ, для встановлення нуклеотидного складу даної НК достатньо визначити кількісне співвідношення основ. Для цієї мети з гидролізатів за допомогою хроматографії на папері або електрофореза (коли в результаті гідролізу одержують нуклеотиди) виділяють індивідуальні з'єднання. Кожна основа незалежно від того, зв'язане воно з вуглеводним фрагментом чи ні, володіє характерним максимумом поглинання в УФ, інтенсивність якого залежить від концентрації. З цієї причини, виходячи з УФ-СПЕКТРІВ виділених з'єднань, можна визначити кількісне співвідношення основ, а отже, і нуклеотидний склад початкової НК. При кількісному визначенні мінорних нуклеотидів, особливо таких нестійких, як дігидроуриділова кислота, користуються ферментативними методами гідролізу (ФДЕ зміїної отрути і селезінки). Використовування описаних вище аналітичних прийомів показало, що НК різного походження полягають за рідкісним виключенням з чотирьох основних нуклеотидів і що зміст мінорних нуклеотидів може мінятися в значних межах. Як буде показано далі, при вивченні нуклеотидного складу ДНК були одержані дані, які допомогли встановити її просторову структуру. 1.2 Значення нуклеїнових кислотЗначення нуклеїнових кислот дуже велике. Особливості їх хімічної будови забезпечують можливість зберігання, перенесення в цитоплазму і передачі по спадку дочірнім кліткам інформації про структуру білкових молекул, як синтезуються в кожній клітці. Білки обумовлюють більшість властивостей і ознак кліток. Зрозуміло тому, що стабільність структури нуклеїнових кислот - найважливіша умова нормальної життєдіяльності кліток і організму в цілому. Будь-які зміни будови нуклеїнових кислот спричиняють за собою зміни структури кліток або активності фізіологічних процесів в них, впливаючи таким чином на життєздатність. Існує два типи нуклеїнових кислот: ДНК і РНК. РНК (рібонуклєїнова кислота), так само як ДНК, є полімером мономерами якого служать нуклеотиди. Азотні підстави ті ж самі, що входять до складу ДНК (аденін, гуанін, цетозін); четверте - урацил - присутній в молекулі РНК замість тіміна. Нуклеотіди РНК містять замість дізоксирібози іншу пентозу - рібозу. ІІ. ДНК2.1 Склад ДНКДНК (дезоксирибонуклеїнова кислота) - біологічний полімер, що складається з двох полінуклеотидних ланцюгів, сполучених один з одним. Мономери, що становлять кожний з ланцюгів ДНК, є складними органічними сполуками, що включають одна з чотирьох азотних основ: аденін (А) або тімін (Т), цитозін (Ц) або гуанін (Г); п'ятиатомний цукор пентозу - дезоксирібозу, на ім'я якої одержала назва і сама ДНК, а також залишок фосфорної кислоти. Ці з'єднання носять назву нуклеотидів. У кожному ланцюзі нуклеотиди з'єднуються шляхом утворення ковалентних зв'язків між дезоксирібозою одного і залишком фосфорної кислоти подальшого нуклеотида. Об'єднуються два ланцюги в одну молекулу за допомогою водневих зв'язків, що виникають між азотними підставами, що входять до складу нуклеотидів, створюючих різні ланцюги. Досліджуючи нуклеотидний склад ДНК різного походження, Чаргафф знайшов наступні закономірності. 1. Всі ДНК незалежно від їх походження містять однакове число пурінових пірімідінових основ. Отже, в будь-якій ДНК на кожен пуріновий нуклеотід доводиться один пірімідіновий. 2. Будь-яка ДНК завжди містить в рівних кількостях попарно аденін тімін, гуанін і цитозін, що звичайно позначають як А=Т і G=C. З цих закономірностей витікає третя. 3. Кількість основ, що містять аміногрупи в положенні 4 пірімідінового ядра і 6 пурінового (цитозін і аденін), рівна кількості основ, що містять оксо-групу в тих же положеннях (гуанін і тімін), тобто A+C=G+T. Ц закономірності одержали назву правил Чаргаффа. Разом з цим було встановлено, що для кожного типу ДНК сумарний зміст гуаніна і цитозіна не рівний сумарному змісту аденіна і тіміна, тобто що (G+C) /(A+T), як правило, відрізняється від одиниці (може бути як більше, так і менше за неї). По цій ознаці розрізняють два основні типи ДНК: А Т-тип з переважним змістом аденіна і тіміна і G C-тип з переважним змістом гуаніна і цитозіна. Величину відношення змісту суми гуаніна і цитозіна до суми змісту аденіна і тіміна, характеризуючу нуклеотидний склад даного виду ДНК, прийнято називати коефіцієнтом специфічності. Кожна ДНК має характерний коефіцієнт специфічності, який може змінюватися в межах від 0,3 до 2,8. При підрахунку коефіцієнта специфічності враховується зміст мінорних основ, а також заміни основних основ їх похідними. Наприклад, при підрахунку коефіцієнта специфічності для ЕДНК зародків пшениці, в якій міститься 6% 5-метілцитозіна, останній входить в суму змісту гуаніна (22,7%) і цитозіна (16,8%). Значення правил Чаргаффа для ДНК стало зрозумілим після встановлення її просторово структури. 2.2 Макромолекулярна структура ДНКУ 1953 р. Уотсон і Крик, спираючись на відомі дані про конформац нуклеозидних залишків, про характер межнуклеотідного зв'язку в ДНК закономірності нуклеотидного складу ДНК (правила Чаргаффа), розшифрували рентгенограми паракрісталічної форми ДНК [так званої В-форми, що утворюється при вогкості вище 80% і при високій концентрації протиіонів (Li+) в зразку]. Згідно х моделі, молекула ДНК є правильною спіраллю, утвореною двома полідезоксирібонуклеотиднимі ланцюгами, закрученими щодо один одного і навколо загальної осі. Діаметр спіралі практично постійний уздовж всієї неї довжини рівний 1,8 нм (18 А).
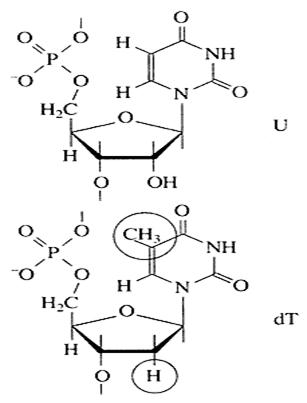
Макромолекулярна структура ДНК. (а) - Модель Уотсона - Крику; (6) - параметры спіралей В-, С - і Т-форм ДНК (проекції перпендикулярно осі спіралі); (в) - поперечний розріз спіралі ДНК у В-форме (заштрихован прямокутники зображають пари основ); (г) - параметри спіралі ДНК в А-форме; (д) - поперечний розріз спіралі ДНК в А-форме. Довжина витка спіралі, який відповідає її періоду ідентичності, склада 3,37 нм (33,7 А). На один виток спіралі доводиться 10 залишків основ в одному ланцюзі. Відстань між площинами основ рівна, таким чином, приблизно 0,34 нм (3,4 А). Площини залишків основ перпендикулярні довгій осі спіралі. Площини вуглеводних залишків декілька відхиляються від цієї осі (спочатку Уотсон і. Крик припустили, що вони паралелі їй). З малюнка видно, що вуглеводофосфатний кістяк молекули обернутий назовні. Спіраль закручена таким чином, що на її поверхні можна виділити дв різні за розмірами борозенки (їх часто називають також жолобками) - велику, шириною приблизно 2,2 нм (22 А), і малу - ширіной близько 1,2 нм (12А). Спіраль - праворучвращаюча. Полідезоксирібонуклеотидні ланцюги в ній антіпаралельні: це означає, що якщо ми рухатимемося уздовж довгої осі спіралі від одного її кінця до іншого, то в одному ланцюзі ми проходитимемо фосфодіефірні зв'язки у напрям 3'à5', а в іншій - у напрямі 5'à3'. Іншими словами, на кожному з кінців лінійної молекули ДНК розташовані 5'-конец однієї і 3'-конец іншого ланцюга. Регулярність спіралі вимагає, щоб проти залишку пурінової основи в одному ланцюзі знаходився залишок пірімідінової основи в іншому ланцюзі. Як вже підкреслювалося, ця вимога реалізується у вигляді принципу утворення комплементарних пар основ, тобто залишкам аденіна і гуаніна в одному ланцюз відповідають залишки тіміна і цитозіна в іншому ланцюзі (і навпаки). Таким чином, послідовність нуклеотидіов в одному ланцюзі молекули ДНК зумовлює нуклеотидну послідовність іншого ланцюга. Цей принцип є головним слідством моделі Уотсона і Крику, оскільки він в дивно простих хімічних термінах пояснює основне функціональне призначення ДНК - бути хранителем генетичної інформації. Закінчуючи розгляд моделі Уотсона і Крику, залишається додати, що сусідні пари залишків основ в ДНК, що знаходиться у В-форме, повернені один щодо одного на 36° (кут між прямими, що сполучають атоми С1' в сусідніх комплементарних парах). ІІІ. РНК3.1 Склад РНКПерші відомості про нуклеотидний склад РНК відносилися до препаратів, що є сумішами клітинних РНК (рибосомних, інформаційних і транспортних) і званим звично сумарною фракцією РНК. Правила Чаргаффа в цьому випадку не дотримуються, хоча певна відповідність між змістом гуаніна і цитозіна, а також аденіна урацила все ж таки має місце. Дані, одержані останніми роками при аналізі індивідуальних РНК, показують, що і на них правила Чаргаффа не розповсюджуються. Проте відмінност в змісті аденіна і урацила, а також гуаніна і цитозіна для більшості РНК невеликі і що, отже, тенденція до виконання вказаних правил все ж таки спостерігається. Цей факт пояснюється особливостями макроструктури РНК. Характерними структурними елементами деяких РНК є мінорні підстави. Відповідн м нуклеотидні залишки звичайно входять до складу транспортних і деяких інших РНК в дуже невеликих кількостях, тому визначення повного нуклеотидного складу таких РНК є іноді вельми складною задачею. 3.2 Макромолекулярна структура РНКХімічно РНК дуже схожа на ДНК. Обидві речовини - це лінійні полімери нуклеотидів. Кожен мономер - нуклеотид - є фосфорилірований N-глікозидом, побудованим із залишку п'ятивуглецевого цукру - пентози, несучого фосфатну групу на гідроксильній групі п'ятого вуглецевого атома (складноефірний зв'язок) азотна основа при першому вуглецевому атомі (N-глікозидний зв'язок). Головна хімічна відмінність між ДНК і РНК полягає у тому, що цукровий залишок мономера РНК - це рібоза, а мономера ДНК - дезоксирібоза, що є похідним рібози, в якому відсутня гідроксильна група при другому вуглецевому атомі (мал.4).
Рис.4. Хімічні формули залишків одного з рібонуклеотидів – уріділово кислоти (U) і гомологичного йому дезоксирібонуклеотида тіміділової кислоти (dT) Азотних основ в РНК чотири види: два пурінових - аденін (А) і гуанін (G) - і два пірімідінових - цитозін (С) і урацил (U) Мономери - рібонуклеотиди РНК - утворюють полімерний ланцюг за допомогою формування фосфодіефірних містків між цукровими залишками (між п'ятим третім атомами вуглецю пентози). Таким чином, полімерний ланцюг РНК може бути представлена як лінійний сахаро-фосфатний кістяк з азотними підставами як бічн групи. Вперше специфічна просторова структура РНК була продемонстрована при розшифровці атомної структури однієї з т-РНК в 1974 р. (мал.5). Згортання полімерного ланцюга т-РНК, що складається з 76 нуклеотидних мономерів, призводить до формування дуже компактного глобулярного ядра, з якого під прямим кутом стирчать два виступи. Вони є короткими подвійними спіралями по типу ДНК, але організовані за рахунок взаємодії ділянок одного і того ж ланцюга РНК. Один з виступів є акцептором амінокислоти і бере участь в синтезі поліпептідного ланцюга білка на рібосомі, а інший призначений для комплементарної взаємодії з кодуючим триплетом (кодоном) т-РНК в тій же рібосоме. Тільки така структура здатна специфічно взаємодіяти з білком-ферментом, що навішує амінокислоту на т-РНК, з рібосомой в процесі трансляції, тобто специфічно "взнаватися" ними.
Мал.5. Атомна (зліва) і скелетна (справа) моделі фенілаланінової т-РНК дріжджів Вивчення ізольованих рібосомних РНК дало наступний разючий приклад формування компактних специфічних структур з ще довших лінійних полімерів цього типу. Рібосома складається з двох нерівних частин - великої і малої рібосомних субчастинок (субодиниць). Кожна субчастинка побудована з одн високополімерної РНК і цілого ряду різноманітних рібосомних білків. Довжина ланцюгів рібосомних РНК вельми значна: так, РНК малої субчастинки бактерійно рібосоми містить більше 1500 нуклеотидів, а РНК великої субчастинки - близько 3000 нуклеотидів. Список використаної літератури1. Н. Грин, У. Стаут, Д. Тейлор – Биология. 2. З.А. Шабарова и А.А. Богданов – Химия нуклеиновых кислот и их полимеров. 3. А.П. Пехов – Биология и общая гинетика. 4. А. Микельсон – Химия нуклеозидов и нуклеотидов. 5. Гауптман, Ю. Грефе, Х. Ремане – Органическая химия. 6. Опарин А.И. Возникновение жизни на Земле (3-е изд). 7. Альтштейн А.Д. Происхождение генетической системы: гипотеза прогенов 8. Б.А. Павлов, А.П. Терентьев "Курс органической химии". |